MECANISMOS IMPLICADOS EN LA FATIGA AGUDA
Gómez-Campos, R.1; Cossio-Bolaños, M.A.2; Brousett Minaya, M.3 y Hochmuller – Fogaca, R.T.4
1Escuela Internacional de Educación Fisica y Deportes – Cuba. Universidad Nacional de San Agustín Arequipa – Perú rossanagomez_c@hotmail.com
2Coordinador de la Escuela de Directores Técnicos de Fútbol. Instituto del Deporte Universitario. Universidad Nacional de San Agustín. Arequipa-Perú. mcossio@udec.cl
3Graduada en Química, Universidad Nacional de San Agustín Arequipa. Universidad Peruana Unión, Lima-Perú. mabromi_1977@hotmail.com
4Jefe del Laboratorio de Fisiología de contracción muscular, UFPR, Curitiba, Brasil fogaca@ufpr.br
Código UNESCO 2411.06 Fisiología del ejercicio
Clasificación del Consejo de Europa: 6. Fisiología del ejercicio.
Recibido 2 de junio de 2009
Aceptado de agosto de 2009
Gómez-Campos, R.; Cossio-Bolaños, M.A.; Brousett Minaya, M. y Hochmuller-Fogaca, R.T. (2010). Mecanismos implicados en la fatiga aguda.Revista Internacional de Medicina y Ciencias de la Actividad Física y el Deporte vol. 10 (40) pp. 537-555.
RESUMEN
La fatiga muscular aguda, puede ser definida como la incapacidad para seguir generando un nivel de fuerza o una intensidad de ejercicio determinada, siendo una situación que se vive, se siente y que atletas o no experimentan; pero debido a su carácter multifactorial, aún los mecanismos de su formación permanecen imprecisos. Se considera la existencia de factores que afectan a nivel muscular, generando la fatiga neuromuscular, y factores que afectan al cerebro, generando la fatiga central. Por lo que el objetivo del presente estudio fue una revisión de literatura, sobre los mecanismos implicados en fatiga aguda, con el propósito de mostrar las diversas vías relacionadas con el proceso de fatiga y dirigir los caminos para su identificación.
PALABRAS CLAVE: Fatiga muscular, mecanismo, músculo.
ABSTRACT
The muscular overtiredness can be defined as the incapacity to raise a level of the muscular strength or a determined intensity of exercise in a situation where athletes experiment efforts. According to the multifactor design of the overtiredness, the mechanisms involved in such a weakened state still remain unclear. With regard to a functional standpoint, this research is dedicated to consider both the central tiredness (nervous system) and the peripheral tiredness (neuromuscular system). The purpose of this study was to review the literature dedicated to the mechanisms involved in the explanation of the overtiredness with regard to various variables needed to be considered.
KEY WORDS: Muscular fatigue, mechanism, muscle.
1. ASPECTOS CONCEPTUALES
La fatiga muscular, es uno de los principales temas investigados dentro del área de fisiología del ejercicio, siendo bien conocida pero no bien definida y entendida (Silva, Goncalves, 2003), por lo que el interés por su estudio ha aumentado. Por mucho tiempo fue descrita como una reacción del músculo al ácido láctico, formulándose la hipótesis de que una cantidad fija de lactato resultarìa en una reducción fija de la tensión (Hill y Kupalov, 1929). Asímismo, Kugelberg & Edstrom (1968), demostraron que las fibras pueden ser altamente fatigables o presentar moderada resistencia a la fatiga, por causa de la depleción de glucògeno en las unidades motoras. De ese modo, son muchos los conceptos dados para definir a la fatiga muscular; fenómeno reversible (Allen, Lamb, Westerblad, 2008), considerado como un conjunto de manifestaciones sintomáticas producidas por trabajo o por un ejercicio prolongado (Rossi, Tirapegui, 1999), o una deficiencia en sustentar un nivel particular de desempeño durante un ejercicio físico (Davis, Bailey, 1997), sea de resistencia o en estados de sobreentrenamiento (Davis, 1995; Jakeman, 1998; Lehmann, et.al 1993; Parry-Billing, et.al 1990, Willians, 1985). Generalmente, también esta asociada a la incapacidad del músculo esquelético de generar elevados niveles de fuerza muscular (Enoka, Stuart, 1992; Green, 1997; Green, 1995) y potencia (Edwards, 1981), a la incapacidad de mantener una determinada intensidad de ejercicio en el tiempo, a la disminución de la velocidad de contracción y al aumento del tiempo de relajaciòn muscular (Allen, Lamergren, Westerblad 1995; Bangsbo, 1997; Davis Bailey, 1997; Mc Kenna, 1992; Newsholme, Blomstron, Ekblom, 1992; Pagala, et.al 1994; Sahlim, 1992, a,b), generando de esa forma una disminución en el rendimiento (Mannion, Dolam, 1996), por lo que, para los entendidos en el área de desempeño físico, resulta un importante indicador de uno de los factores de riesgo para la ocurrencia de lesiones por sobrecarga (Ascensao, et.al 2003), mostrándose como un antecedente de algún tipo de injuria relacionada al deporte (Duarte, Dias, Melo 2008).
1. ETIOLOGÌA DE LA FATIGA
Los mecanismos etiológicos responsables de la fatiga, han recibido importante atención de varios fisiologistas y bioquímicos por más de un siglo (Fitts, 1994), existiendo una gran cantidad de estudios en la búsqueda cada vez más pormenorizada de su etiología, en la perspectiva de la mejora del desempeño en el deporte de alto rendimiento (Allen, Langergrem, Westerblod, 1995; Bangsbo 1997; McKenna, 1992; Nicol, Koni, Marconnet, 1991; Noakes, 2000; Sahlim, 1992 a,b), así como los trabajos realizados en el ámbito de la recuperación funcional de los sujetos con patologías o lesiones en determinadas estructuras del sistema nervioso (Castro, et.al 2000; Lindeman, et.al 1999; Svantesson, et.al 1999) y las investigaciones en sujetos con patologías neuromusculares (Drost, et.al 2001, Kent-Braun, Miller 2000; Pagala, et.al 1993; Sunnerhagen, et.al 2000), le confieren ser considerada uno de los temas más estudiados en fisiología del ejercicio.
Se han sugerido algunas causas que dan origen a la fatiga muscular, como las alteraciones del pH, de la temperatura y del flujo sanguíneo, la acumulación de productos del metabolismo celular (especialmente de los que resultan de la hidrólisis del ATP, como el ADP, AMP, IMP, Pi y amonio), la pérdida de la homeostásis del ión Ca2+, el papel de la cinética de algunos iones en los medios intra y extracelular (como el K+, Na+, Cl– Mg2+), la lesión muscular (inducida por el ejercicio) y el stress oxidativo (Ascensao, et.al, 2003), como se muestran en la figura 1. Sin embargo, a pesar de la gran cantidad de estudios, los mecanismos asociados a su etiología se encuentran aún por determinar (Green, 1995; Mc Lester, 1997), consecuentemente una de las principales dificultades al investigar la fatiga se debe a la naturaleza multifactorial y complejidad de la misma (McArdle, Katch y Katch, 1998; Kirkendall, 2000), de donde se deriva una división funcional de la misma, tal como se muestra en la figura 2, en fatiga central y fatiga periférica (Duarte, Dias, Melo, 2008), que lleva en cuenta factores metabólicos interactivos que afectan a los músculos (fatiga periférica) y al cerebro (fatiga central) durante la realización del trabajo físico intenso en atletas y otros individuos (Lehmann, Foster & Keul, 1993).
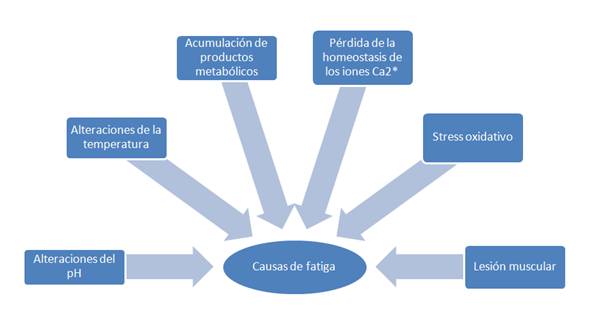
Figura 1. Causas sugeridas de fatiga. Elaborado por el autor.
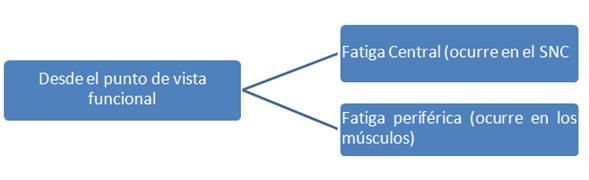
Figura 2 Clasificación de la fatiga. Elaborado por los autores.
2. FATIGA PERIFÉRICA O NEUROMUSCULAR
Cuando el músculo recibe un estímulo en forma de potencial de acción, el retículo sarcoplasmático (RS) libera Ca+, el cual se liga a la troponina y forma el complejo tropomiosina, exponiendo a su vez la actina, el sitio de ligación para la cabeza de la miosina (Duarte, Dias, Melo, 2008), asì la cabeza de miosina desligada de la actina, en presencia de ATP, utiliza la energía de la hidrólisis del ATP para moverse, ligándose enseguida a la actina, empujando el filamento fino a lo largo del filamento grueso, haciendo que el sarcómero se acorte y se produzca el proceso de contracción muscular (Scott, et.al, 2001).
La incapacidad de mantener ese potencial de acción (que depende de la capacidad de recapturar los iones de potasio K+, para dentro de la célula y expeler los iones sodio Na+, con el fin de repolarizar la membrana sarcoplasmática y permitir la entrada de un nuevo impulso eléctrico, Green,1997), constituye un importante factor desencadenador de la fatiga; de ese modo, se considera que la Fatiga Periférica, resulta de las alteraciones de la homeostasis en el músculo esquelético (Ascensao, et.al 2003), debido a una falla o limitación de uno o más procesos en la unidad motora (nervios periféricos, ligaciones neuromuscular o fibras musculare) (Edwards, 1981). Así, como consecuencia de la disminución del pH cistólico, se produce un aumento de la capacidad del RS para retener el Ca+, reduciendo la estimulación del proceso contráctil (Nakamura, Schwartz, 1972). En ese sentido, varios estudios (Allen, et.al 1992; Appell, Soares & Duarte 1992, Gandevia, 1992; Mc Kenna, 1992; Mc Lester, Junior 1997; Sahlim 1992), le dan un sustento científico a la fatiga muscular, enfocado en los factores que resultan de la disfunción del proceso de contracción, como impedimentos en la transmisión neuromuscular en el RS; así también, otro estudio (Willians, 1998), realizado en modelos animales, corrobora que el desarrollo de la fatiga esta asociado a alteraciones funcionales del RS y a las propiedades del aparato contráctil, relacionadas a los puentes cruzados del ciclo cinético con el aumento de Ca+; asimismo Allen, Lannergren, Westerblad (1995), demostraron en un estudio, en fibras musculares intactas aisladas, que durante la fatiga se da una disminución del Ca2+ con consecuente disminución de la fuerza.
La fatiga muscular, depende del tipo, duración e intensidad del ejercicio, del tipo de fibra muscular reclutada, del nivel de entrenamiento del sujeto y de las condiciones ambientales de realización del ejercicio (Davis, Fitts, 2001; Enoka, Stuart, 1992; Fitts, Metzger, 1988; Robert, Smith, 1989. En ese sentido, cabe considerar que en el músculo esquelético, la glucosa es el almacén de glucógeno, y este es la fuente de mayor almacenamiento de energía durante varias formas de actividad muscular (Allen, Lamb, Westerblad, 2008), por lo que, durante la realización de ejercicios físicos se producen alteraciones metabólicas de suma importancia (Coyle, et.al 1983; Katz, et.al 1991), que envuelven la deficiencia de energía para el trabajo muscular, conocida también como “Hipótesis de la depleción de glucógeno” (Snyder, 1988), donde de acuerdo con la duración e intensidad del ejercicio hay una activación de los sistemas energéticos y metabólicos específicos (Lancha Junior, 1996) generando la disminución en la disponibilidad de substratos energéticos al músculo esquelético activo durante el ejercicio (Ascensao, et.al 2003). Esta hipótesis que fue justificada por Davis, Fitts (2001), Fitts, Metzger (1998), Sahlim (1992, 1996, 1998), indicando que, durante la realización de ejercicios de alta intensidad, donde hay una producción elevada de energía obtenida a través de la vía glucolítica anaeróbica, se genera una disminución del glucógeno muscular, con la consiguiente producción de lactato y la intervención del sistema de regeneración del ATP, Creatinfosfato. En el caso de la ejecución de ejercicios prolongados de baja intensidad, donde existe una baja producción de energía, el ATP se obtiene de la degradación oxidativa de substratos metabólicos como el glucógeno muscular, la glucosa sérica, los ácidos grasos libres oriundos de los músculos o del tejido adiposo (Duarte, Dias, Melo, 2008), siendo característica de la fibra muscular, el consumo de ATP, y producción de ADP y Pi, mucho más rápido de lo que se regenera (Allen, Lamb, Westerblad, 2008).
La importancia del glucógeno muscular en ejercicios de resistencia, ha sido reconocida desde la década de los 60 (Hermansan, et.al, 1967; Bergstrom, et.al, 1967), como es demostrado en los estudios de Pernow y Saltim (1971), donde concluyen que durante la ejecución de ejercicios de larga duración se producen cambios en la utilización de substratos por el músculo, los cuales pueden ocasionar efectos secundarios en la manutención de los niveles plasmáticos de substratos y hormonios a nivel del metabolismo intracelular del glucógeno en el músculo, pero aún la posible relación entre la depleción de ese substrato y la fatiga permanece imprecisa (Duarte, Dias, Melo, 2008), no pudiendo despreciar su participación en el proceso de fatiga, la que puede ocurrir por vía indirecta, pues la disminución del glucógeno muscular puede comprometer el acoplamiento, excitación-contracción (Chin y Allen, 1997; Stephenson, et.al 1999).
Por otro lado, durante el ejercicio moderado, el músculo esquelético activo se torna la principal fuente de amonio (Lowesntein, Goodman, 1978), que es producido por las reacciones celulares durante el ejercicio asociado tanto a la fatiga central como la periférica (Banister & Cameron, 1990; Banister, Rajendra & Mutch, 1985; Guezennec, et.al 1998). Durante el ejercicio prolongado la concentración plasmática de amonio puede elevarse significativamente, lo que depende de la intensidad y duración del ejercicio (Eriksson.et.al, 1985; Lo y Dudley 1987; Ament, et.al, 1997; Snow, et.al, 2000), y aunque parte de la cantidad de amonio permanece en el musculo esquelético la mayor parte es liberada a la circulación sanguínea, que al ser capaz de atravesar la barrera hematoencefálica (Bachmann, 2002), puede acumularse en altos niveles en los espacios intra y extracelulares del SNC y ocasionar importantes efectos a nivel de los neurotransmisores en el metabolismo cerebral y en la circulación (Banister y Cameron, 1990), cuyos disturbios pueden contribuir negativamente en la funciones del cuerpo durante el ejercicio (Butterworth, et.al 1988; Davis y Bailey 1997), provocando perturbaciones cerebrales que pueden influir en el desarrollo de la fatiga central. Sin embargo, una reducción en los niveles plasmáticos de amonio durante el ejercicio, puede aumentar la capacidad individual para soportar la intensidad del ejercicio exhaustivo (Yvan et.al 2002; Nybo, et.al 2005).
Otro factor habitualmente discutido, como posible causa de la fatiga, es el acumulo de lactato o acidosis metabólica, inducida por el ejercicio de alta intensidad y de corta duración (Wagenmakers, Coacley, Edwards, 1990), donde se obtiene energía de modo predominantemente anaeróbico, y el aumento de la concentración de iones H+, ocasiona la disminución del pH (producto de la disociación del ácido láctico), factor que está asociado a la inhibición de la enzima PKF (fosfofrutoquinasa) y reducción en la Glucólisis (Rossi, Tirapegui, 1999) pudiendo desencadenar la acidez dentro de la célula y ser letal para la misma o contribuir con el proceso de fatiga precoz (Sahlin, 1992), así como lo muestra el estudio realizado por Sahlin, Palmskog, Hultman (1978), en un sprint experimental, donde el pH intramuscular puede disminuir de su valor de reposo (7,0) hasta 6,4 una vez que el pH sanguíneo varía de 7,4 a 6,8-6,9 respectivamente. Sin embargo, el músculo puede realizar contracciones a alta potencia con elevadas concentraciones de lactato, desde que el pH se mantenga próximo a 7,0 (Santos, Dezan, Sarraf, 2003); pero si fuera inferior a 7,0 (donde hay acumulo de H+) se verificará una disminución de la potencia muscular; por lo que existen varios estudios (Balog, Fitts, 2001; Chin, Allen, 1998; Dutka, Lamb, 2000; Favero, et.al, 1997; Posterino, Fryer, 2000; Stienen, Papp, Zaremba, 1999; Thompson, Fitts, 1992), que determinan la influencia del aumento de la concentración del H+ en el proceso de contracción muscular y consecuente desarrollo de la fuerza. Con todo, aún la literatura muestra controversias en cuanto a la existencia de una relación directa entre la disminución del pH intracelular y la disminución de la fuerza muscular, así como la influencia de los iones lactato e H+ en la fatiga muscular (Roberts, Smith, 1989).
Cabe resaltar, que Bangsbo, et.al (1996), refiere que la disminución del pH, no se presenta como la única causa de la fatiga en ejercicios de intensidad elevada y de corta duración, atribuyendo que el acumulo de K+ intersticial también tienen un papel importante en el desarrollo de la fatiga, como fue corroborado en los estudios de Cooke, Pate (1990), Degror, et.al (1991), McLester, (1997), Thompson, Fitts (1994), que concluyen que la presencia de la fatiga se debe en mayor medida, a la concentración de H2PO4 (forma protonada del Pi) que del pH, siendo sugerido por Fitts, Metzger (1988), que la influencia del H+ en la disminución de la fuerza, se puede deber al consecuente aumento de las concentraciones del H2PO4.
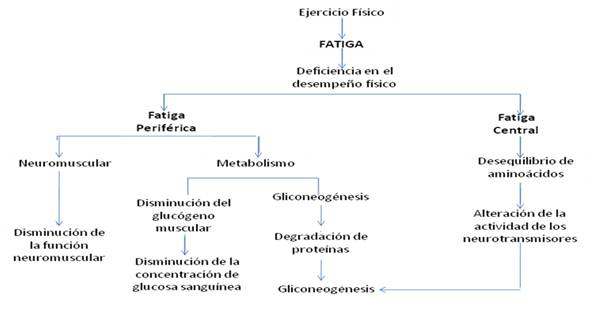
Figura 3. Factores relacionados a la Fatiga. Adaptado de Rossi Tirapegui (1999)
3. FATIGA CENTRAL
La fatiga central es probablemente la que presenta mayores controversias entre los investigadores (Gandevia, Allen, Mckenzie, 1994), al referirse a las alteraciones en el funcionamiento cerebral (Davis, Bailey, 1997), traducidas en una falla voluntaria o involuntaria en la conducción del impulso (Stackhouse, et.al 2000; Sunnerhagen, et.al 2000), que pueden ocurrir en uno o más niveles de las estructuras nerviosas que intervienen en la actividad física, lo cual puede provocar una alteración en la transmisión desde el SNC o en el reclutamiento de los axones motores (Santos, Dezan, Sarraf, 2003).
El papel del SNC en el origen de la fatiga, investigado a nivel muscular por Allen, Lannergren, Westerblad (1995), Stackhouse, et.al (2000), determinan que la fuerza máxima que el sujeto consigue generar voluntariamente, es comparada con la fuerza producida supramaximalmente por electroestimulación exógena del nervio motor o del propio músculo (Allen, Lannergren, Westerblad, 1995; Davis, Fitts, 2001), concluyéndose inicialmente que la disminución de la actividad nerviosa, y por ende del Sistema Nervioso (SN), no representaba un factor relevante en la instalación de la fatiga muscular (Ascensao, et.al, 2003), sin embargo, estudios recientes justifican la importancia de los mecanismos centrales en la manutención de un determinado nivel de fuerza (Davis, Bailey, 1997; Davis, Fitts, 2001; Gandevia, 1992).
Por bastante tiempo, el papel de las proteínas y aminoácidos en la actividad física, no fue considerado relevante, asimismo, aún en los últimos 30 años, los estudios se concentraban en el efecto del ejercicio sobre el metabolismo de carbohidratos y grasas, siendo las proteínas ampliamente ignoradas (Lemon, 1997); por lo que a partir de los años 70 y 80, es que el interés por conocer los efectos del ejercicio sobre el metabolismo de proteínas y aminoácidos ha aumentado considerablemente, así como lo muestran varios estudios (Applegate & Grivetti, 1997; Banister & Cameron, 1990; Dohm, Beecher, Warren & Willians, 1981; Lancha Junior, 1996; Marquesi & Lancha Junior, 1997; Wu, 1998), afirmando que los aminoácidos contribuyen significativamente durante el ejercicio prolongado e consecuentemente sobre el rendimiento humano.
Asimismo, siendo los aminoácidos, precursores de algunos neurotransmisores (Chaouloff, 1989; Dishman, 1997), cabe destacar, que uno de los probables mecanismos asociados a la fatiga central, esta relacionado a la alteraciones en la síntesis y en la actividad de algunos neurotransmisores (Silva, Oliveira, Silva, 2006), hecho de donde deriva la llamada “Hipótesis de la Fatiga Central”. Asimismo, las aminas biogénicas, son compuestos que poseen como grupos funcionales las aminas, formadas en su mayoría a partir de la descarboxilación de los aminoácidos aromáticos (fenilalanina, tirosina e triptófano) histidina y sus derivados (Lenhinger, 1989), que se encargan de regular el metabolismo de los mamíferos (Rossi, Tirapegui, 1999). Así, la serotonina (5-hidroxitriptamina: 5-HT) derivada del triptófano, la histamina deriva de la histidina y las catecolaminas – dopamina, norepinefrina y epinefrina derivan de la tirosina. En consecuencia, producto del ejercicio físico intenso y prolongado, estas aminas o neurotransmisores influyen en el desarrollo de la fatiga, aunque aún poco se sabe respecto a los mecanismos que envuelven este proceso de fatiga (Bailey Davis & Ahlborn, 1992; Davis & Bailey, 1997).
De igual manera, ha sido observado un aumento en las concentraciones de triptófano libre, consecuentemente de la serotonina, en ejercicios de larga duración (Silva, Oliveira, Silva, 2006), donde existe una mayor estimulación de la lipólisis y por ende aumento en la concentración plasmática de ácidos grasos, que se unen a la albúmina y contribuyen para el aumento de las concentraciones de Triptófano libre (TRF libre); bajo esta forma es transportado a través de la barrera hematoencefalica (Blomstrand, 2001, Knaflitz, Bonato, 1999; Newsholme, Blomstrand, Ekblom, 1992); por lo que, se relaciona con el sistema de fatiga muscular, debido a la posible alteración que pueda ocasionar en la percepción del esfuerzo muscular (Newsholme, Acworth, 1987), por su función en la regulación del ciclo circadiano (Terrados, Fernandez, 1997), indisposición, somnolencia, falta de atención (Davis & Bailey, 1997), el humor (Lieberman, et.al 1985) y supresión del apetito (Blundell, 1992; Lyons & Truswell, 1988; Wellman, 1992; Weltzin, et.al 1994), por todo lo anterior, es que la serotonina es uno de los neurotransmisores más estudiados (Chauoloff, 1989; Chauoloff, et.al 1985; Chauoloff, Laude & Elghozi, 1989; Newsholme &, Blomstrand , 1996).
Por otro lado, la dopamina (DA), fue el primer neurotransmisor en ser estudiado en la fatiga central (Rossi, Tirapegui, 1999), asociándolo a aspectos como la locomoción, emoción y aprendizaje (Barenoud, 2000), existiendo varios estudios como el realizado en pacientes con Parkinson, donde se verificó una mejoría en el control motor después del tratamiento con l-dopa (precursor de la DA) (Berne, Levy, 2000); así también, en el ambiente deportivo las investigaciones de Bhagat & Wheeler (1973), Clarkson & Thompson (1997), Wellman (1992), en que unos atletas, al ingerir drogas (anfetamina), sufrieron un aumento en la función dopaminérgica del cerebro, conllevó a aumentar su desempeño, siendo uno de los mecanismos posibles de la DA, que explican su asociación a la habilidad de está, para inhibir parte de la síntesis y metabolismo de la 5HT retardando el tiempo para la fatiga central (Kirkendall, 2000). De igual manera fueron observadas, las consecuencias de la administración de algunos aminoácidos de cadena ramificada (AACR) (leucina, isoleucina y valina), en cuanto ser inhibidores del aumento de la síntesis cerebral de serotonina (Blomstrand, 2001; Davis, 1995; Kreider, Miriel, Bertun, 1993; Newsholme, Blomstrand, Ekblom, 1992; Varnier, et.al1994)
Finalmente, otro neurotransmisor relacionado con la fatiga central es la acetilcolina (Santos, Dezon, Sarraf, 2003), donde la tasa de síntesis de acetilcolina es determinada por la disponibilidad de su precursor, la colina (Ascensao, et.al, 2003), pudiendo contribuir en el metabolismo de la fatiga central (Kirkendall, 2000) y aunque no sea definitiva su asociación a la fatiga central o periférica, la restricción en el consumo de colina puede disminuir la velocidad de transmisión de los impulsos en el músculo esquelético (Xia, 2000).
4. CONSIDERACIONES FINALES
Queda evidente, la existencia de una gran fuerza científica dirigida al estudio de los mecanismos de la fatiga central y periférica, que intentan relacionar la causa, o probablemente las causas, de la fatiga del músculo esquelético, entendiéndola no sólo como un factor o una regulación, sino también como una suma de factores y circunstancias; y aún entre los investigadores, es unánime la idea de que en esa área, hay que tener mucho cuidado al reunir las informaciones sobre un determinado aspecto, por lo que, nos lleva a concluir que aún se tenga una gran cantidad de datos sobre diversos aspectos de la fatiga, no existe aún una definición final sobre las causas reales de la fatiga, confortándonos el hecho de que de las investigaciones en este tema están surgiendo de forma exponencial y aún tendremos más informaciones y datos por reunir.
REFERENCIAS BIBLIOGRAFICAS
Allen, D.; Lamb, G.; Westerblad, H. (2008) Skeletal Muscle Fatigue: Cellular Mechanics. Physilo Rev 88:287-332.
Allen, D.; Lännergren, J.; Westerblad H. (1995) Muscle cell function during prolonged activity: cellular mechanisms of fatigue. Experimental Physiology 80, p.497-527.
Allen, D.; Westerblad, H.; Lee, J.; Lannergren, J. (1992) Role of excitation-contraction coupling in muscle fatigue. Sports Medicine, v.13, n.2, p.116-26.
Ament, W.; Huizenga, J.; Mook, G.; Gip. H.; Verkerke, G. (1997) Lactate and ammonia concentration in blood and sweat during incremental cycles, Cicloergometer exercise. Int J Sports Med; v.18, n.1, p. 35-39.
Appell, H.; Soares, J.; Duarte, J. (1992) Exercise, muscle damage and fatigue. Sports Medicine, v.13, n.2, p.108-15.
Applegate, E.; Grivetti, L. (1997) Search for the competitive edge: a history of dietary fads and supplements. Journal of Nutrition; v.127, n.5, p.869S-73S.
Ascensão, A.; Magalhães, J.; Oliveira, J.; Duarte, J.; Soares, J. (2003) Fisiologia da fadiga muscular. Delimitação conceptual, modelos de estudo e mecanismos de fadiga de origem central e periférica. Revista Portuguesa de Ciências do Desporto; vol. 3, n.1, p.108-123.
Bachmann, C. (2002) Mechanisms of hyperammonemia. Clin Chem Lab Med. v. 40, n. 7, p. 653-662.
Balog, E.; Fitts, R. (2001). Effects of depolarization and low intracellular ph on movement currents of frog skeletal muscle fibers. J Appl Physiol 90 228-234
Bailey, S.; Davis, J.; Ahlborn, E. (1992) Effect of increased brain serotonergic activity on endurance performance in the rat. Acta Physiologica Scandinavica, v.145, p.75-6.
Bangsbo, J.; Madsen, K.; Kiens, B.; Richter, E. (1996). Effect of muscle acidity on muscle metabolism and fatigue during intense exercise in man. J Physiol 495 (2): 587-596
Bangsbo, J. (1997). Physiology of muscle fatigue during intense exercise. In T Reilly, M Orme, The clinical pharmacology of sport and exercise, Elsevier Science BV, 123-130.
Banister, E.W.; Cameron, B.J.C. (1990) Exercise induced hyperammonemia: peripheral and central effects. International Journal of Sports Medicine, v.11, n.2, p.S129-42, 1990.
Banister, E.; Rajendra, W.; Mutch, B. (1985). Ammonia as an indicator of exercise stress implications of recents findings to sports medicine. Sports Medicine, v.2, n.1, p.34-46.
Barenoud, P. et al. (2000) Evaluation of simple and complex sensorimotor behaviors in rats with a partial lesion of the dopaminergicnigrostriatal system. Eur J Neurosci, 12: 322-336.
Bergstrom, J.; Hermansen, L.; Hultman, E.; Saltin, B. (1967) Diet, muscle glycogen and physical performance. Acta Physiol Scand, v. 71, n. 2, p. 140- 150.
Berne, R.; Levy, M. (2000) Fisiologia. 4. ed. Guanabara.
Bhagat, B.; Wheeler, N. (1973) Effect of amphetamine on the swimming endurance of rats. Neuropharmacology; v. 12, p. 711- 3.
Butterworth, R.; Girard, G.; Giguere, J. (1988) Regional differences in the capacity for ammonia removal by brain following portocaval anastomosis. J Neurochem, v. 51, n. 2, p. 489-490.
Blundell, J. (1992) Serotonin and the biology of feeding. American Journal of Clinical Nutrition, v.55, p.155S-9S.
Blomstrand, E. (2001). Amino acids and central fatigue. Amino Acids 20 25-34
Castro, M.; Aplle, D.; Melton-Rogers, S.; Dudley, G. (2000). Muscle fiber type-specific myofibrillar Ca2+ ATPase activity after spinal cord injury. Muscle and Nerve 23 119-121.
Chaouloff, F. (1989) Physical exercise and brain monoamines: a review. Acta Physiologica Scandinavica, v.137, p.1-13.
Chaouloff, F.; Elghozi, J.; Guezennec, Y.; Laude, D. (1985) Effects of conditioned running on plasma, liver and brain tryptophan and on brain 5- hydroxytryptamine metabolism of the rat. British Journal of Pharmacology, v.86, p.33-41.
Chaouloff, F.; Laude, D.; Elghozi, J. (1989). Physical exercise: evidence for differential consequences of tryptophan on 5-HT synthesis and metabolism in central serotonergic cell bodies and terminals. Journal Neural Transmission, v.78, p.121-30.
Chin, E.; Allen, D. (1997) Effects of reduced muscle glycogen concentration on force, Ca2+ release and contractile protein function in intact mouse skeletal muscle. J Physiol. v. 498, p. 17-29.
Chin, E.; Allen, D. (1998). The contribution of ph-dependent mechanisms to fatigue at different intensities in mammalian single muscle fibers. J Physiol 512 (3): 831-840
Clarkson, P.; Thompson, H. (1997) Drugs and sport: research findings and limitations. Sports Medicine, v.24, n.6, p.366-84.
Coyle, E.; Hagberg, J.; Hurley, B., Martin, W., Ehsani, A., Holloszy, J. (1983) Carbohydrate feeding during prolonged strenuous exercise can delay fatigue. J Appl Physiol. v. 55, p. 230-235.
Cooke, R.; Pate, E. (1990) The inhibition of muscle contraction by the products of ATP hydrolysis. In Taylor, A.; Gollnick, P.; Green, H.; Ianuzzo, C.; Noble, Métivier, G.; Sutton, J. Biochemestry of Exercise VII, Champaign, IL: Human Kinetics, 59-72
Davis, M.; Fitts, R. (2001) Mechanisms of muscular fatigue. In P Darcey, ACSM´S resource manual – guidelines for exercise testing and prescription, Baltimore: Lippincott Williams & Wilkins, 184-190
Davis, M.; Bailey, S. (1997) Possible mechanisms of central nervous system fatigue during exercise. Med Sci Sports Exerc 29 (1): 45-57.
Davis, J. (1995a) Carbohydrates, branched-chain amino acids, and endurance: the central fatigue hypothesis. International Journal of Sport Nutrition, v.5, p.S29-38.
Degroot, M., Massie, M., Boska, M., Gober, J., Miller, R., Dishman, R. (1997) Brain monoamines exercise and behavioral stress: animal models. Medicine and Science in Sports and Exercise, v.29, n.1, p.63-74.
Dishman, R.K. (1997) Brain monoamines, exercise, and behavioral stress: animal models. Medicine and Science in Sports and Exercise, Madison, v.29, n.1, p.63-74.
Dohm, G.; Beecher, G.; Warren, R.; Williams, R. (1981) Influence of exercise on free amino acid concentrations in rat tissues. Journal of Applied Physiology, v.50, n.1, p.41-4.
Duarte, V.; Dias, D. & Melo, H. (2008) Mecanismo Moleculares da fadiga. Brazilian Journal of Biomotricity. V.2, n.1,p.3-38.
Dutka, T.; Lamb, G. (2000) Effect of lactate on depolarization- induced Ca2+ release in mechanically skinned muscle fibers. Am J Physiol 278 C517-C525.
Drost, G.; Blok, J.; Stegman, D.; Van Dick, J.; Van Egelen, B.; Zwarts, M. (2001) Propagation disturbance of motor unit action potentials during transient paresis in generalized myotonia – A high-density surface EMG study. Brain 124 352-360.
Enoka, R.; Stuart, D. (1992) Neurobiology of muscle fatigue. J Apll Physiol 72 (5): 1631-1648.
Edwards, R. (1981) Human muscle function and fatigue. Londres. Edic. Whelan; 82:1-18.
Eriksson, L.; Broberg, S.; Bjorkman, O.; Wahren, J. (1985) Ammonia metabolism during exercise in man. Clin Physiol. v. 5, n. 4, p. 325-336.
Favero, T.; Zable, A.; Colter, D.; Abramson J. (1997) Lactate inhibits Ca2+-activated Ca2+-channel activity from skeletal muscle sarcoplasmatic reticulum. J Appl Physiol 82 (2): 447-452.
Fitts, R. (1994) Cellular mechanisms of muscle fatigue. Physiol Rev. v. 74, n. 1, p. 49-94.
Fitts, R.; Metzger, J. (1998) Mechanisms of muscular fatigue. In J Poortmans, Principals of Exercise Biochemistry, Basel: Krager, 212-229.
Gandevia, S. (1992) Some central and peripheral factors affecting human motoneuronal output in neuromuscular fatigue. Sports Medicine, v.13, n.2, p.93-8.
Gandevia, S.; Allen, G.; Mckenzie. (1994) Central fadigue: Critical issues, quantification and pratical applications. In: Gandevia SC, ENOKA RM, Mccomas AJ et al. Fatigue: Neural and muscular mechanisms. Advances in Experimental Medicine and Biology; 384:281-294.
Green, H. (1997) Mechanisms of muscle fatigue in intense exercise. J Sports Sci 15 247-256.
Green, S. (1995) Measurement of anaerobic work capacities in humans. Sports Med 19 (1): 32-42.
Guezennec, C.; Abdelmalki, A.; Serrurier, B.; Merino, D.; Bigard, X.; Berthelot, M.; Pierard, C.; Peres, M. (1998) Effects of prolonged exercise on brain ammonia and amino acids. International Journal of Sports Medicine, v.19, p.323-7.
Hermansen, L.; Hultman, E.; Saltin, B. (1967) Muscle glycogen during prolonged severe exercise. Acta Physiol Scand. v. 71, n. 2, p. 129-139.
Hill, A.;Kupalov, P. (1929) Anaerobic and Aerobic Activity in Isolated Muscle Proc. R. Soc. v. 105, n. 737, p. 313-328.
Jakeman, P. (1998) Amino acid metabolism, branchedchain amino acid feeding and brain monoamine function. Proceedings of the Nutrition Society, v.57, p. 35-41.
Katz, A.; Raz, I.; Spencer, M.; Rising, R.; Mott, D. (1991) Hyperglycemia induces accumulation of glucose in human skeletal muscle. Am J Physiol. v. 33 260, n. 4, p. R698-703.
Kent-Braun, J.; Miller, R. (2000) Central fatigue during isometric exercise in amytrophic lateral sclerosis. Muscle and Nerve 23 909-914
Kugelberg, E.; Edstrom L. (1968) Differential histochemical effect of muscle contrations on phosphorylase and glycogen in various types of fibres: Relation to fatigue. J. Neurol Neurosurg Psychiatry. 31:415-23.
Knaflitz, M.; Bonato, P. (1999) Time-frequency methods applied to muscle fatigue assessement during dynamic contractions. J Electromyogr Kinesiol 9 337-350.
Kirkendall, D. (2000) Fatigue from voluntary motor activitiy. In: Exercise and sport science. Lippincott Willians & Wilkin; p.97-104.
Kreider, R.; Miriel, V.; Bertun, E. (1993) Amino acid supplementation and exercise performance – Analysis of proposed ergogenic value. Sports Med 16 ( 3): 190-209
Lancha Junior, A. (1996) Atividade física, suplementação nutricional de aminoácidos e resistência periférica à insulina. Revista Paulista de Educação Física, v.10, n.1, p.68-75.
Lehmann, M.; Foster, C.; Keul, J. (1993) Overtraining in endurance athletes: a brief review. Medicine and Science in Sports and Exercise, v.25, n.7, p.854- 62.
Lehninger, A. (1989) Princípios de bioquímica. 5.ed. São Paulo, Sarvier.
Lemon, PWR. (1997) Influência da proteína alimentar e do total de energia ingerida no aumento da força muscular. Sports Science Exchange, 10.
Lieberman, H.R.; Corkin, S.; Spring, B.J.; Wurtman, R.J.; Growdon, J.H. (1985) The effects of dietary neurotransmitter precursors on human behavior. American Journal of Clinical Nutrition, v. 42, p. 336- 70.
Lindeman, E.; Spaans, F.; Reulen, J.; Leffers, P.; Drukker, J. (1999) Progressive resistance training in neuromuscular patients. Effects on force and surface EMG. J Electromyogr Kinesiol 9 (6): 379-384.
Lo, P.; Dudley, G. (1987) Endurance training reduces the magnitude of exercise-induced hyperammonemia in humans. J Appl Physiol; v. 62, n. 3, p. 1227-1230.
Lowesntein, J.M.; Goodman, M.N. (1978) The purine nucleotide cycle in skeletal muscle. Federation Proceedings, v.37, p.2308-12.
Lyons, P.M.; Truswell, A.S. (1988) Seretonin precursor influenced by type of carbohydrate meal in healthy adults. American Journal of Clinical Nutrition, v.47, p.433-9.
Mannion, A.F.; Dolan, P. (1996) Relationship between myoelectric and mechanical manifestations of fatigue in the quadriceps femoris muscle group. Eur.J.Appl Physiol.Occup. Physiol; 74(5): 411-419.
Marquesi, M.L.; Lancha Junior, A.H. (1997) Possível efeito da suplementação de aminoácidos de cadeia ramificada, aspartato e asparagina sobre o limiar aeróbico. Revista Paulista de Educação Física, v.11, n.1, p.90-101.
McArdle, W.; Katch, F.; Katch, V. (1998) Fisiologia do exercicio: energia, nutrição e desempenho humano. 3.ed. Rio de Janeiro, Guanabara Koogan.
McLester, J (1997). Muscle contraction and fatigue – the role of adenosine 5´-diphosphate and inorganic phosphate. Sports Med 23 (5): 287-305
McKenna, M. (1992) The roles of ionic processes in muscular fatigue during intense exercise. Sports Med 13 (2): 134-145.
Nakamura, Y.; Schwartz, A. (1972) The influence of hydrogen ion concentrations on calcium binding and release by skeletal muscle sarcoplasmic reticulum. J Gen Physiol; 59:22-32.
Newsholme, E.; Blomstrand, E.; Ekblom, B. (1992) Physical and mental fatigue: Metabolic mechanisms and importance of plasma amino acids. Sports Med 48 (3): 477-495.
Newsholme, E.; Acworth, E. (1987) Amino acids, brain neurotransmitters and a functional link between muscle and brain that is important in susteained exercise. In: Benzi G, ed. Advances in myochemistry. London Libbey Eurotext; 127-133.
Newsholme, E.; Blomstrand, E. (1996) The plasma level of some amino acids and physical and mental fatigue. Experientia, v.52, p.413-5.
Nicol, C.; Komi, P.; Marconnet, P. (1991) Fatigue effects of marathon running on neuromuscular performance, II – Changes in force, integrated electromyographic activity and endurance capacity. Scand J Med Sci Sports 1: 18-24
Noakes, TD. (2000). Physiological models to understand exercise fatigue and the adaptations that predict or enhance athletic performance. Scand J Med Sci sports 10 (3): 123-145.
Nybo, L.; Dalsgaard, M.; Steensberg, A.; Moller, K.; Secher, N. (2005) Cerebral ammonia uptake and accumulation during prolonged exercise in humans. J Physiol. v. 563, p. 285–290.
Pagala, M.; Ravindran, K.; Amaladevi, B.; Namba, T.; Grob, D. (1994) Potassium and caffeine contractures of mouse muscles before and after fatiguing stimulation. Muscle and Nerve 17: 852-859.
Pagala, M.; Nandakumar, N.; Venkatachari, S.; Ravindran, K.; Amaladevi, B.; Namba, T.; Grob, D. (1993) Mechanisms of fatigue in normal intercostal muscle and muscle from patients with myastenia gravis. Muscle and Nerve 16: 911-921
Parry-Billings, M.; Blomstrand, E.; McAndrew, N.; Newsholme, E. (1990). A communicational link between skeletal muscle, brain, and cells of the immune system. International Journal of Sports Medicine; v.11, n.2, p.S122-8.
Pernow, B.; Saltin, B. (1971) Availability of substrates and capacity for prolonged heavy exercise in man. J App Physiol. v. 31, p. 416-422.
Posterino, G.; Fryer, M. (2000) Effects of high myoplasmatic L-lactate concentration on E-C coupling in mammalian skeletal muscle. J Appl Physiol 89: 517-528
Rossi, L.; Tirapegui, J. (1999) Aspectos atuais sobre o exercicio físico, fadiga e nutricao. Rev. Paul. Educ. Fís., São Paulo, 13(1): 67-82.
Roberts, D.; Smith, D. (1989) Biochemical aspects of peripheral muscle fatigue – a review. Sports Med 7: 125-138
Sahlin, K. (1992a) Metabolic factors in fatigue. Sports Med 13 (2): 99-107.
Sahlin, K. (1992b) Metabolic aspects of fatigue in human skeletal muscle. In P Marconnet, P Komi, B Saltin, O Sejested, Muscle Fatigue in Exercise and Training, Basel: Karger, 54-68.
Sahlin, K. (1996) Energy metabolism and muscle fatigue during exercise. In Steinacker, Ward, The physiology and pathophysiology of exercise tolerance, New York: Plenum Press, 45-51
Sahlin, K.; Tonkonogi, M.; Söderlund, K. (1998) Energy supply and muscle fatigue in humans. Acta Scand Physiol 162 261-266
Sahlin, K.; Palmskog, G.; Hultman, E. (1978) Adenine nucleotide and IMP contents of the quadriceps muscle in man after exercise. Pflugers Archiv; 374:193-198.
Santos, M.; Dezan, V.; Sarraf, T. (2003) Bases metabólicas da fadiga muscular aguda. Rev. Bras. Ciên. e Mov. Brasília v. 11 n. 1 p. 07-12.
Silva, S.; Goncalves, M. (2003) Análise da fadiga muscular pela amplitude do sinal eletromiográfico. Rev Bras. Ci. E Mov. Brasilia. V.11 n.3. p.15-20.
Stephenson, D.; Nguyen, L.; Stephnson, G. (1999) Glycogen content and excitation-contaction coupling in mechanically skinned muscle fibres of the cane toad. J Physiol. v. 519, n. 177-187.
Scott, W.; Stevens, J.; Binder-Macleod, S. (2001) Human Skeletal Muscle Fiber TypeHuman Skeletal Muscle Fiber Type Classifications. Physical Therapy. v. 81, p. 1810-1816.
Silva, A.; Oliveira, F.; Silva, M. (2006) Mecanismo de Fadiga durante o exercicio fìsico. Revista Brasileira de cineantropometria Desempenho Human; 8(1):105-113.
Snyder, A. (1998). Overtraining and glycogen depletion hypothesis. Medicine and Science in Sports and Exercise, v.30, n.7, p.1146-50.
Snow, R.; Carey, M.; Stahis, C.; Febbraio, M.; Hargreaves, M. (2000) Effect of carbohydrate ingestion on ammonia metabolism during exercise in humans. J Appl Physiol. v. 88, p. 1576-1580.
Stienen, G.; Papp, Z.; Zaremba, R. (1999) Influence of inorganic phosphate and ph on sarcoplasmatic reticular ATPase in skinned muscle fibers of Xenopus laevis. J Physiol 518 (3): 735-744.
Svantesson, U.; Sunnerhagen, K.; Carlsson, U.; Grimby, G. (1999) Development of fatigue during repeated eccentricconcentric muscle contractions of plantar flexors in patients with stroke. Arch Phys Med Rehabil 80: 1247-1252.
Stackhouse, S.; Dean, J.; Lee, S.; Binder-Macload, S. (2000) Measurment of central activation failure of the quadriceps femoris in healthy adults. Muscle and Nerve 23: 1706-1712.
Sunnerhagen, K.; Carlsson, U.; Sandberg, A.; Stålberg, E.; Hedberg, M.; Grimby, G. (2000) Electrophysiologic evaluation of muscle fatigue development and recovery in late polio. Arch Phys Med Rehabil 81: 770-776.
Terrados, N.; Fernandez, B. (1997) Fatiga muscular. In: Fatiga muscular en el rendimento deportivo. Sintesis.
Thompson, L., Fitts, R. (1992), Muscle fatigue in the frog semitendinosus: role of the high-energy phosphates and Pi. Am J Physiol 263 (Cell Physiol 32): C803-C809,
Varnier, M.; Sarto, P.; Martines, D.; Lora, L.; Carmignoto, F.; Leese, G.; Naccarato, R. (1994) Effect of infusing branchedchain amino acid during incremental exercise with reduced muscle glycogen content. Eur J Appl Physiol 69: 26-31
Wagenmakers, A.; Coacley, J.; Edwards, R. (1990). Metabolism of branched-chain amino acids and ammonia during exercise: clues from McArdle’s disease. Int J Sports Med; 260: 883-890.
Wellman, P. (1992) Overview of adrenergic anorectic agents. American Journal of Clinical Nutrition, v.55, p.193S-8S.
Weltzin, T.; Fernstrom, J.; McConaha, C.; Kaye, W. (1994) Acude tryptophan depletion in bulimia: effects on large neutral amino acids. Biological Psychiatry, v.14, n.35, p.388-97.
Williams, C. (1985) Nutritional aspects of exercise-induced fatigue. Proceedings of the Nutrition Society, v.44, p.245-56.
Williams, J.; et al. (1998) Functional aspects of skeletal muscle contractile apparatus and sarcoplasmic reticulum after fatigue. American physiological society; 619-626.
Yuan, Y.; So, R.; Wong, S.; Chan, K. (2002) Ammonia threshold- comparation to lactate threshold, correlation to other physiological parameters and response to training. Scand J Med Sci Sports. v. 12, p. 358-364.
Wu, G. (1998) Intestinal mucosal amino acid catabolism. Journal of Nutrition, v.128, n.8, p.1249-52,.
Xia, N. (2000) Effects of dietary choline levels on human muscle function. MS tesis, Boston University College of Engineering, 1991 Apud Kirkendall DT. Fatigue from voluntary motor activitiy. In: Exercise and sport science. Garrett WE, Kirkendall DT. Lippincott Willians & Wilkins.