SIGNIFICADO FISIOLÓGICO DE LA TRANSICIÓN AERÓBICA-ANAERÓBICA
Calderón, F.J.1; Benito, PJ.2; Peinado, A.B.3 y Díaz, V.4
1(franciscojavier.calderon@upm.es) Doctor en Medicina y Cirugía. Profesor Titular de Universidad, Facultad de Ciencias de la Actividad Física y del Deporte – INEF. Universidad Politécnica de Madrid, España.
2(pedroj.benito@upm.es) Doctor en Ciencias de la Actividad Física y del Deporte. Profesor Titular Interino, Facultad de Ciencias de la Actividad Física y del Deporte – INEF. Universidad Politécnica de Madrid, España.
3(anabelen.peinado@upm.es) Licenciada en Ciencias de la Actividad Física y del Deporte. Becaria de doctorado, Facultad de Ciencias de la Actividad Física y del Deporte – INEF. Universidad Politécnica de Madrid, España.
4(victor.diaz@upm.es) Licenciado en Ciencias de la Actividad Física y del Deporte. Técnico de Laboratorio, Facultad de Ciencias de la Actividad Física y del Deporte – INEF. Universidad Politécnica de Madrid, España.
RECIBIDO 2 de octubre 2008
ACEPTADO 11 de noviembre de 2008
CLASIFICACIÓN UNESCO 2411
Calderón, F.J.; Benito, PJ.; Peinado, A.B. y Díaz, V. (2008). Significado fisiológico de la transición aeróbica-anaeróbica. Revista Internacional de Medicina y Ciencias de la Actividad Física y el Deporte vol. 8 (32) pp. 321-337.
RESUMEN
La transición aeróbica-anaeróbica ha sido muy estudiada, proponiéndose diversos métodos para su determinación. Estos procedimientos se pueden dividir en ergoespirométricos y no ergoespirométricos. Los primeros se basan en modificaciones de parámetros del intercambio respiratorio, obteniéndose el umbral anaeróbico ventilatorio (UAventilatorio). Los segundos se sustentan en los cambios producidos por diversos parámetros determinados en sangre, músculo u otras secreciones (saliva). Con estos procedimientos se obtienen los siguientes umbrales anaeróbicos: UAláctico, UAcatecolaminas, UAEMG y UAsaliva.
A pesar de la vasta información relativa a este fenómeno, desconocemos cuál es el mecanismo fisiológico que permite explicar la coincidencia en el tiempo de los diferentes métodos de determinar el UA. Consideramos que el UA constituye para el sistema nervioso central (SNC) una situación de estrés de todo el organismo. Así, el SNC ordena un aumento de actividad nerviosa de forma generalizada a través de sus dos subdivisiones (somático y vegetativo). La activación simultánea de ambas subdivisiones justificaría la coincidencia en el tiempo de métodos de determinación del UA tan diversos y aparentemente no relacionados. Desconocemos cuál es la información procesada y cuáles son las estructuras del SNC que intervienen. Probablemente, la propia señal eferente propuesta para explicar el significado fisiológico del UA, sea también la información que recibe el SNC. El hecho de que aumente, por ejemplo, la concentración de lactato a partir del UA, constituiría una información para el SNC sobre la situación del organismo próxima al límite.
PALABRAS CLAVE: umbral anaeróbico, sistema nervioso central, ventilación, lactato.
ABSTRACT
The aerobic-anaerobic transition has been studied along years. Several methodologies have been proposed: ergoespirometric and non ergoespirometric. The first ones are based on modifications of parameters of the gas exchange, obtaining the ventilatory threshold. The next are based in the changes produces by several parameters measured in blood, muscle or other secretions (saliva). With these procedures the following anaerobic thresholds are obtained: lactic, catecholamines, electromiography (EMG) and saliva.
In spite of the great amount of information regarding this phenomenon, we do not know which is the physiological mechanism that allows to explain the coincidence in the time of the different methodologies to determine the anaerobic threshold. We think that the anaerobic threshold is an alarm signal for the central nervous system (CNS). The simultaneous activation of the subdivisions of the CNS (somatic and vegetative) would justify the coincidence in the time of the different anaerobic thresholds. We do not know which is the processed information and which are the structures of the CNS that takes part. Probably, the own efferent signal propose to explain the physiological meaning of the anaerobic threshold, is also the information that receives the CNS. The increases of the lactate concentration, for example, would warn the CNS that the organism is near the limit.
KEY WORDS: anaerobic threshold, central nervous system, ventilation, lactate.
1. INTRODUCCIÓN
Cualquier persona ha experimentado que subir 5 pisos determina que su corazón vaya más deprisa (aumento de la frecuencia cardiaca (FC)) y su respiración se acelere (incremento de la frecuencia respiratoria (FR)). Esto es lo más evidente, aunque además se realicen otros ajustes más difíciles de percibir, tales como el incremento del volumen sistólico (VS) o del volumen alveolar (Va). El aumento de la FC y FR constituyen las respuestas del sistema cardiovascular y aparato respiratorio, respectivamente. Si cualquier persona sube todos los días 3 o 4 veces los cinco pisos percibirá, al cabo del tiempo, que su corazón no late tan deprisa y que su respiración es más lenta. Es decir, el entrenamiento habrá desencadenado la adaptación del corazón y del aparato respiratorio. Este ejemplo tan simple sirve para ilustrar la respuesta y adaptación al ejercicio dinámico.
A lo largo de la historia de la Fisiología del Ejercicio se han desarrollado diferentes procedimientos para valorar la respuesta y adaptación del organismo (1, 2). La ergoespirometría constituye un método estandarizado y utilizado de forma habitual en los laboratorios de Fisiología del Ejercicio (3). La información aportada por esta prueba es, en realidad, un compendio de la respuesta integrada del organismo al ejercicio dinámico, pues permite conocer el funcionamiento del sistema cardiovascular, aparato respiratorio y de forma indirecta, del metabolismo (4, 5). De los parámetros obtenidos en una prueba de esfuerzo, los más importantes, desde un punto de vista práctico son: el consumo de oxígeno máximo (VO2 máx.) y la transición aeróbica anaeróbica. La importancia de estos dos parámetros radica en dos hechos: 1º) desde el punto de vista teórico, para intentar explicar el funcionamiento del organismo en condiciones límite; 2º) desde el punto de vista práctico, debido a que muchos de los fundamentos del entrenamiento se basan en conocer estos dos parámetros.
Fisiológicamente, el VO2 máx. constituye el parámetro “central” de una prueba ergoespirométrica, pues es el más representativo del funcionamiento global del organismo, ya que integra la función de múltiples aparatos y sistemas (sistema cardiovascular, aparato respiratorio, transporte sanguíneo de los gases y metabolismo). Cuando un deportista se encuentra en una situación real, intenta llevar una intensidad de ejercicio lo más próxima a su VO2 máx. Por lo tanto, adquiere especial relevancia medir cuánto es capaz el organismo de acercarse al límite del rendimiento, intentando aguantar el máximo tiempo posible. Sin embargo, parece natural pensar que el “límite fisiológico” no se pueda mantener durante mucho tiempo. Por este motivo, a lo largo de los años ha adquirido gran importancia conocer cuánto tiempo se puede sostener un ejercicio dinámico lo más próximo al VO2 máx. Como la capacidad para mantener un ejercicio prolongado se relaciona estrechamente con el metabolismo, los investigadores pensaron que el “limite” se relacionaba con el momento en el que el metabolismo pasaba de una situación aeróbica a anaeróbica. A este momento se le conoce como transición aeróbica-anaeróbica (2, 6-8). Aunque el término utilizado de forma más habitual es el de umbral anaeróbico, desde el punto de vista de concepción del fenómeno, es preferible denominarlo transición aeróbica-anaeróbica. El término umbral supone un valor bien delimitado, como si fuera el dintel de una puerta, mientras que el término transición sugiere un camino, que se acerca más a la realidad. Además, la existencia de “dos umbrales” es un argumento más para que se entienda como zona y no como umbral. No obstante, a efectos prácticos y de simplificación, en esta revisión adoptaremos el término genérico de umbral anaeróbico (UA) para referirnos al segundo umbral ventilatorio (VT2).
La importancia del UA ha determinado que, durante muchos años, el esfuerzo de los investigadores haya ido dirigido a proponer métodos para su determinación (2, 6, 9). En la actualidad, los procedimientos incruentos para la determinación del UA se encuentran considerablemente estandarizados, de manera que en los sistemas automatizados se han desarrollado programas informáticos específicos. Sin embargo, paradójicamente, como el momento de “cambio metabólico” es un fenómeno fisiológicamente muy complejo, los procedimientos también lo son. El objetivo de este trabajo es ofrecer una visión pedagógica de este fenómeno fisiológico tan complejo, realizando una revisión de la literatura científica publicada al respecto.
2. EXPLICACIÓN FISIOLÓGICA DE LOS PROCEDIMIENTOS DE DETERMINACIÓN DE LA TRANSICIÓN AERÓBICA-ANAERÓBICA
Los métodos o procedimientos de determinación del UA se pueden dividir en: 1) métodos ergoespirométricos y 2) métodos no ergoespirométricos. Los primeros son referidos a los que se obtienen mediante diferentes parámetros del intercambio gaseoso. Los segundos son los que valoran el UA a través de diferentes parámetros analizados en sangre, músculo u otros líquidos orgánicos. Aunque existe controversia, es necesario tener presente que los dos métodos están fisiológicamente relacionados. Algunos autores mantienen que los parámetros ergoespirométricos se relacionan estrechamente con parámetros metabólicos o de otra índole (8). Por el contrario, Brooks (1985) opina que son fenómenos aislados aunque relacionados (10). El objetivo de este epígrafe es dar una explicación fisiológica global e integrada de todos los métodos destinados a detectar el UA.
2.1. Relación fisiológica entre los diferentes procedimientos de determinación del UA.
2.1.1. Umbral anaeróbico ventilatorio (UAventilatorio)
Diferentes parámetros del intercambio gaseoso han sido empleados para determinar los umbrales ventilatorios (9). De forma general, se determinan cuando las variables de intercambio respiratorio experimentan una modificación en su respuesta. El problema de los diferentes métodos estriba en el considerable componente de subjetividad. Al ser métodos visuales, es un aspecto importante la manera en la que el investigador “observa” las variaciones que se producen en el comportamiento de los diferentes parámetros. Para eliminar dicho componente de subjetividad, algunos autores han propuesto procedimientos matemáticos (11, 12) o bien la combinación de diferentes métodos (13).
Todos los parámetros del intercambio gaseoso pulmonar empleados en la determinación del UA (ventilación, equivalentes respiratorios para el oxígeno y para el dióxido de carbono, etc.) están directamente relacionados con los mecanismos de regulación de la respiración. Teniendo en cuenta que el control respiratorio tiene lugar, de forma general, en el encéfalo, el significado funcional de los cambios producidos en las variables señaladas a partir del UA es doble: 1. Constituiría la “señal eferente” enviada por los centros de regulación de la respiración, como resultado de detectar una situación de alarma para la homeostasis. 2. Sería la “señal aferente” enviada por diversos receptores (pulmonares y extrapulmonares), para que los centros de regulación de la respiración respondieran de “forma exagerada”.
2.1.2. Umbral anaeróbico láctico (UAláctico)
Como el organismo responde ante el estrés que supone el ejercicio físico de forma integrada, muchos investigadores han encontrado relaciones entre el UA determinado por ergoespirometría y la “tendencia” de muchos otros parámetros. En plasma, el parámetro que más se ha estudiado ha sido el lactato. De hecho, para algunos investigadores es el patrón oro con el que se comparan otros métodos, aunque ha sido y es origen de gran controversia. Las razones de la controversia, relacionadas entre sí, son:1. De carácter metodológico, pues se requiere la extracción de sangre y la determinación de la concentración de lactato. 2. De carácter fisiológico, pues la concentración de lactato es el resultado de la relación producción/eliminación (14).
Atendiendo a las características histoquímicas y bioquímicas de las fibras musculares, el aumento de la producción de lactato se explica cuando se produce una mayor activación de las fibras musculares tipo II. Teniendo en cuenta los conceptos de unidad motora y orden de reclutamiento, la activación de las fibras tipo II tendría lugar de forma preferente a partir del UA. El aumento del lactato en plasma influye en la modificación del estado ácido-base. Así, a partir del UA, se produce un aumento de la concentración de protones ([H+]), así como variaciones en los parámetros fisico-químicos que determinan el estado ácido base. Algunos autores han propuesto que se puede determinar el UA mediante parámetros del estado ácido-base, para conocer el momento de la transición metabólica (15, 16).
2.1.3. Umbral anaeróbico electromiográfico (UAEMG)
En razón a lo expuesto, algunos investigadores han intentado, mediante métodos incruentos (EMG: electromiografía de superficie), detectar la mayor actividad de las fibras tipo II. La EMG ha sido utilizada para determinar la frecuencia de estimulación de las unidades motoras. Moritani y col. (1982) y posteriormente otros investigadores (17-22) que han utilizado esta técnica, han demostrado un cambio en la actividad electromiográfica integrada durante la transición aeróbica-anaeróbica. Al valor umbral detectado mediante EMG se le ha denominado umbral electromiográfico.
2.1.4. Umbral anaeróbico de catecolaminas (UAcatecolaminas)
Algunos investigadores han propuesto que a partir del UA se produce un incremento desproporcionado de la concentración de catecolaminas en sangre (23-25). Además, este aumento se relaciona estrechamente con otros procedimientos para la determinación del UA. Al valor a partir del cual se produce un aumento de la concentración de catecolaminas en plasma se le denomina umbral de catecolaminas.
2.1.5. Umbral anaeróbico de saliva (UAsaliva).
También se ha estudiado el comportamiento de la concentración de iones o moléculas orgánicas en determinados líquidos corporales en el momento de la transición aeróbica-anaeróbica. Concretamente, las variaciones de lactato u otros iones o moléculas observadas en la saliva, ha sido un método propuesto por diferentes investigadores (26-29). La base fisiológica del UA mediante la composición de la saliva no es inédita, pues ya en 1963, Salminen y Kontinen describieron las modificaciones de la misma durante el ejercicio (30). Se ha demostrado que existe una buena correlación entre el lactato plasmático y el de la saliva, y entre la variación de la concentración de cloro ([Cl–]) y sodio ([Na+]) y los parámetros del intercambio gaseoso.
2.1.6. Umbral anaeróbico de la frecuencia cardiaca (UAFC)
Conconi y col. han sostenido que la relación lineal frecuencia cardiaca (FC)/intensidad no presenta la misma pendiente a lo largo de una prueba de esfuerzo incremental (31, 32). El punto donde se produce un cambio de la pendiente se relaciona, según estos autores, con el UA. Desde el punto de vista del control del entrenamiento, las expectativas que este método abrió fueron considerables, pues bastaría con registrar la FC en una prueba de campo. Sin embargo, existe controversia en relación al método (31-37).
El UA determinado mediante la FC tiene cierta coherencia fisiológica. Parece lógico pensar que, en un momento dado, los centros nerviosos para el control cardiovascular desencadenen una respuesta distinta de la actividad vegetativa sobre el nódulo marcapasos. El resultado sería el cambio de pendiente en la relación FC/intensidad a partir del UA. A diferencia de la frecuencia respiratoria que aumenta desproporcionadamente, la FC sigue aumentando hasta el máximo, pero en menor magnitud. La razón puede ser el hecho de que el aumento de la FC condicione la duración del ciclo cardiaco, principalmente la diástole. Así, un incremento mayor de la FC a partir del UA podría comprometer el llenado ventricular, de manera que los centros de regulación cardiovascular disminuyen su actividad sobre el nodo sinusal.
2.2. Significado fisiológico de los métodos de determinación del UA
De forma general, la respuesta de las variables ergoespirométricas presenta dos estadios o fases, que se pueden denominar como: lineal y no lineal. La fase lineal representa la proporcionalidad del organismo ante el estrés que supone el ejercicio. Por ejemplo, a mayor demanda más elevados son el gasto cardiaco y la ventilación alveolar. Por el contrario, la fase denominada no lineal implica que la respuesta del organismo es más elevada que las necesidades. Por ejemplo, a partir del UA, la ventilación es mayor que el aprovechamiento que se puede realizar.
La linealidad del VO2 y de la producción de dióxido de carbono (VCO2) se explica por el aumento de actividad del aparato respiratorio, sistema cardiovascular y capacidad de transporte de los gases respiratorios. Durante la fase de linealidad, el organismo suministra el oxígeno suficiente a las mitocondrias del tejido muscular, de manera que éstas pueden oxidar el combustible, principalmente carbohidratos. Al mismo tiempo, extrae el dióxido de carbono que se elimina a la atmósfera. La relación de intercambio gaseoso (CR) se encuentra próxima a la unidad.
El cambio o modificación de la respuesta lineal a partir del UA, como se ha señalado anteriormente, se relaciona estrechamente con la modificación del metabolismo. En términos sencillos, se puede decir que la respuesta no lineal representa el paso del metabolismo aeróbico al anaeróbico: el organismo no aporta el oxígeno suficiente al músculo. Así, parte del piruvato no puede oxidarse en las mitocondrias y se reduce a lactato, evitando que el potencial redox del citosol aumente de forma considerable. Al mismo tiempo, los sistemas de eliminación comienzan a perder eficiencia en relación a la producción. La respuesta no lineal de las diferentes variables ergoespirométricas se podría justificar en base a los mecanismos de regulación cardiovascular y respiratoria.
¿Cuál es el mecanismo fisiológico que permite explicar la aparente coincidencia en el tiempo de los diferentes métodos de determinación del UA? De forma elemental, el UA constituye para el sistema nervioso central (SNC) una situación de estrés, al igual que para el organismo en su conjunto. Así, el SNC “ordena” un aumento de actividad que es enviada a través del sistema nervioso periférico (SNP). Como los nervios espinales contienen fibras nerviosas tanto del sistema nervioso somático como del sistema nervioso vegetativo, el resultado es una actividad simultánea de ambas subdivisiones del SNP. Por lo tanto, la mayor actividad de ambos sistemas justificaría (figura 1): la activación de las unidades motoras FT (fast twich), que se traduciría en el UAEMG y secundariamente en la mayor producción de lactato (UAláctico); la mayor actividad de los centros de regulación de la respiración, que se traduciría en el UAventilatorio; la activación vegetativa generalizada que sería responsable del UAcatecolaminas y del UAsaliva y la menor activación de los centros de regulación cardiovascular, que se traduciría en el UAFC.
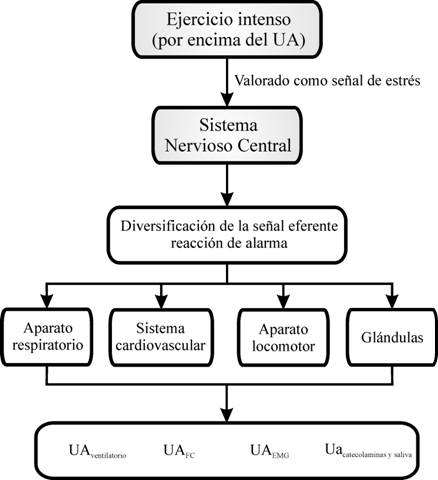
Figura 1. Mecanismo fisiológico propuesto para explicar la aparente coincidencia en el tiempo de los diferentes métodos de determinación del UA.
El sistema nervioso respondería al estrés que supone el ejercicio intenso mediante una doble activación: córtico-adrenal y simpático-adrenal. Sin embargo, esta visión integrada es de compleja comprobación experimental. Algunas de las preguntas que surgen al respecto son: ¿cómo se produce el aumento de la activación simpático-adrenal a partir del UA?, ¿qué estructuras del SNC intervienen?, ¿qué tipo de información procesa el SNC para “modificar” la respuesta a partir del UA?, etc. Alguna de estas preguntas podría ser contestada, en parte, en función del conocimiento fisiológico que se tiene del concepto de estrés. Por lo tanto, esta visión integrada del UA se limita a señalar que únicamente conocemos la “señal de salida”, que se traduce en una coincidencia en el tiempo del UAventilatorio, UAláctico, UAEMG, UAcatecolaminas y UAsaliva.
3. MECANISMO INTEGRADOR DE LA RESPUESTA DEL ORGANISMO
La diversificación de la señal eferente es, probablemente, la forma de poner en alerta a la totalidad del organismo. En términos de Seyle, la respuesta del organismo a partir del UA es inespecífica, como sucede en otras formas de estrés. Esta respuesta se ha mantenido a lo largo de la evolución. Se desconocen las señales que recibe el SNC a la hora de elaborar una respuesta múltiple y qué centros nerviosos elaboran, dirigen y coordinan una señal eferente tan compleja.
3.1. ¿Qué estructuras del SNC intervienen durante el ejercicio intenso? ¿Qué tipo de información procesa el SNC para modificar la respuesta a partir del UA?
El hecho de que a partir del UAEMG se sugiera una activación de las unidades motoras tipo FT, puede hacer pensar que la médula espinal pudiera ser un centro nervioso de elaboración de la respuesta. Asimismo, el hecho de que aumente la ventilación (UAventilatorio) puede ser debido a que los centros bulbares que intervienen en la regulación de la respiración, elaboren la respuesta. Finalmente, la respuesta del organismo al ejercicio intenso es llevada a cabo por los sistemas simpático-adrenal e hipofisario-adrenal, de manera que los centros nerviosos que coordinan ambos podrían ser los responsables del UAcatecolaminas y del UAsaliva. Es decir, siguiendo un orden caudo-craneal, podríamos pensar que los centros nerviosos de elaboración estarían situados en las estructuras nerviosas derivadas del mesencéfalo primitivo. Los centros nerviosos del tronco encefálico y médula espinal estarían subordinados a los anteriores.
Por otra parte, para explicar la activación de las unidades motoras tipo FT (UAEMG) que permitirían dar respuesta al aumento de la concentración de lactato (UAláctico), es necesario que de forma simultánea a la activación hipofisaria-adrenal y simpático-adrenal, se produzca un cambio en el orden de reclutamiento, a través de las vías motoras descendentes que actúan sobre la vía final común. Los sistemas eferentes de los centros supraespinales, al actuar sobre las motoneuronas espinales, desencadenan la excitación de las unidades motoras de diferente umbral, de manera que pueden estimular a las de mayor valor (unidades FT) e inhibir las del menor valor (unidades ST – slow twich).
Además, a nivel de la médula espinal también se puede ejercer control sobre el reclutamiento. Las características electrofisiológicas de las motoneuronas del asta anterior permiten una activación secuencial de las mismas.El principio del tamaño determina que exista una relación inversa entre la resistencia de la membrana de las motoneuronas y el tamaño del soma. También, se ha sugerido que las poblaciones de unidades motoras pueden activarse de forma diferente, siguiendo órdenes de reclutamiento distintas. Esto permitiría la activación de toda una población de unidades motoras y evitar la saturación de unidades con un bajo nivel de activación, permitiendo que respondieran rápidamente a las influencias espinales y supraespinales. Por lo tanto, el sistema motor integrado por los centros de elaboración, vías nerviosas y unidades motoras, se encargaría de enviar la señal eferente necesaria, que coincidiría con el UAEMG y el UAláctico.
Mayor dificultad entraña la segunda pregunta. Al ser la señal eferente tan extensa, parece lógico pensar que, igualmente, las señales aferentes a los centros nerviosos sean también múltiples. Es muy probable que la información a los centros nerviosos que coordinan la respuesta al ejercicio proceda de: receptores cinestésicos, receptores del sistema cardiovascular y receptores del aparato respiratorio (38-40).
En resumen, la activación simultánea de los sistemas que se encargan de la coordinación vegetativa (hipofisario-adrenal y simpático-adrenal) y del sistema motor, justificaría la coincidencia en el tiempo de métodos de determinación del UA tan diversos y aparentemente no relacionados (UAventilatorio, UAláctico, UAEMG, UAcatecolaminas y UAsaliva).
3.2. Sentido fisiológico del mecanismo integrador
Es probable que a partir de cierta intensidad la información que llega al SNC sirva para que interprete que todos los órganos y sistemas se encuentran muy próximos a su límite máximo. Por ejemplo, consideremos uno de los parámetros más comúnmente utilizados para conocer que se ha llegado al máximo: la FC. Aunque puede ser objeto de debate, una forma muy sencilla de conocer el límite máximo cardiaco es restarle a 220 la edad de una persona. Al valor obtenido se le denomina FC máxima teórica. Teniendo en cuenta que el UA se alcanza alrededor del 80 % de la FC máxima teórica, siendo el margen para alcanzar el máximo un 20 %. Al igual que otras señales de alarma, como la sed o el hambre, el organismo debe conocer cuánto margen tiene antes de que peligre su homeostasis (41-45).
Por lo tanto, probablemente las diferentes señales enviadas por el SNC durante el UA sirven, además, como señal de retroalimentación. Así, el aumento de la concentración de lactato, catecolaminas y el incremento ventilatorio, por ejemplo, constituirían señales para que el SNC conozca que se está alcanzando el máximo. Como el límite para realizar ejercicio intenso es multifactorial, parece coherente que la información para detectarlo también lo sea. De forma general, la limitación para realizar ejercicio puede venir condicionada por: una incapacidad del sistema cardiovascular y aparato respiratorio para suministrar el oxígeno y eliminar el dióxido de carbono en relación a la demanda metabólica de los tejidos; y una incapacidad de utilización de los sustratos energéticos. Ambas condiciones se encuentran íntimamente ligadas y quedan reflejadas en la figura 2. Aunque es discutible cuál es la función fisiológica limitante para realizar ejercicio máximo, parece que el responsable puede ser el sistema cardiovascular.
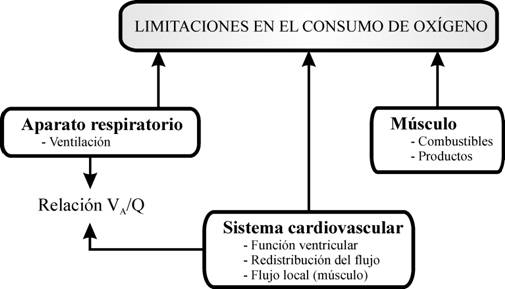
Figura 2. Limitantes del consumo de oxígeno.
Si el UA es considerado como una señal de alarma para la integridad del organismo, ¿cambia con el entrenamiento?Durante muchos años, los entrenadores han depositado muchas expectativas en la importancia práctica del UA. Se pensaba que, en personas muy entrenadas, el VO2 máx. apenas se modificaba y que los resultados en la mejora del rendimiento se podían explicar por la modificación del UA. En nuestra opinión, en personas muy entrenadas, el UA es tan poco modificable como el VO2 máx. La razón estriba, precisamente, en la consideración del UA como señal de alarma. Mucho ha sido el esfuerzo dedicado a conocer las modificaciones del VO2 máx. durante el proceso entrenamiento, pero más limitados han sido los estudios encaminados a conocer el desplazamiento del UA con este proceso. Las diferencias encontradas para el VO2 máx. oscilan entre el 4 y el 33 %, debido a: estado previo de entrenamiento, programa de entrenamiento (intensidad, duración, frecuencia y método) y herencia (46).
En la tabla 1 se presentan los diferentes estudios que han analizado la evolución de la transición aeróbica-anaeróbica, que experimenta una oscilación considerable cuando este proceso se encuentra al inicio (umbral ventilatorio 1: VT1) y un menor porcentaje de adaptación cuando el metabolismo adquiere un predominio anaeróbico (umbral ventilatorio 2: VT2). No obstante, en la mayor parte de los estudios las diferencias no eran significativas. Como el nivel de resistencia de los deportistas estudiados era muy variable, las diferencias del UA también lo eran. Así, las diferencias para el VT1 oscilan entre el 0,5 y el 22 % y las diferencias para el VT2 son del 2,5 al 12,8 %. También se observa que cuanto más entrenada es la población objeto de estudio, menor es el desplazamiento del UA (46).
|
I |
II |
III |
IV |
||
|
Berg, K (47) |
Umbral láctico (mmol·L-1) |
8,1 ± 0,4 |
8,2 ± 0,3 |
||
|
Zapico, AG (ciclistas) (48) |
Umbral láctico (mmol·L-1) |
11,3 ± 1,1 |
12 ± 1,1 |
11 ± 1 |
|
|
VT1 (% VO2 máx.) |
52 ± 2,1 |
60 ± 1,8 |
62 ± 1,7 |
||
|
VT2 (% VO2 máx.) |
82,5 ± 1,7 |
87,9 ± 1 |
90,1 ± 1,4 |
||
|
Zapico, AG (triatletas) (49) |
Umbral láctico (mmol·L-1) |
12,1 ± 1,4 |
10,3 ± 1,4 |
14,1 ± 1,3 |
|
|
VT1 (% VO2 máx.) |
50 ± 2,5 |
61 ± 2,1 |
56 ± 2 |
||
|
VT2 (% VO2 máx.) |
83,2 ± 2 |
83,2 ± 1,2 |
86,2 ± 1,6 |
||
|
Koutedakis, Y (50) |
VT2 (% VO2 máx.) |
85,4 ± 5,2 |
82,2 ± 5,2 |
76,9 ± 4,2 |
|
|
Pardo, FJ (51) |
Umbral láctico (mmol·L-1) |
7,9 ± 0,5 |
8,1 ± 0,5 |
6,7 ± 0,5 |
|
|
VT1 (% VO2 máx.) |
71,6 ± 1,3 |
72 ± 1,5 |
73,1 ± 1,3 |
||
|
VT2 (% VO2 máx.) |
88,7 ± 0,8 |
86,7 ± 1,1 |
89,2 ± 1,1 |
||
|
Svedenhag, J (52) |
VT2 (% VO2 máx.) |
85,9 ± 1,3 |
85,6 ± 1,3 |
82,8 ± 1,1 |
85,2 ± 1,2 |
|
Tanaka, K (53) |
VT1 (% VO2 máx.) |
75,1 ± 5,5 |
74,5 ± 5,2 |
74,5 ± 3,5 |
|
|
White, JA (54) |
VT1 (W) |
281 ± 29,9 |
317 ± 29,1 |
||
|
VT2 (W) |
518 ± 37,5 |
557 ± 38,5 |
|||
|
Yuan, Y (55) |
VT1 (% VO2 máx.) |
62,2 ± 5,6 |
56,9 ± 7,2 |
63,5 ± 4,7 |
|
|
VT2 (% VO2 máx.) |
76 ± 5,9 |
76 ± 4,8 |
78,6 ± 6,4 |
||
Tabla 1. Parámetros de la transición aeróbica-anaeróbica (umbrales ventilatorios o umbral láctico) a lo largo de una temporada.
Parece lógico pensar que cuando un deportista se encuentre en el momento óptimo de forma, sus variables fisiológicas se encuentren al límite de la mejora. ¿Cuál puede ser la razón o razones que expliquen la estabilización del UA? Al ser considerado como una señal de alarma, las variaciones que pueda experimentar están sujetas a muchos parámetros difíciles de controlar, referidos al mecanismo fisiológico del estrés. Además, los diferentes métodos son muy rudimentarios para determinar ligeros matices de adaptación en personas muy entrenadas.
4. CONCLUSIONES
Con lo visto hasta ahora, considerar el UA como una señal de alarma parece lógico si tenemos en cuenta la respuesta integrada del organismo y no de sus partes de forma aislada. Aunque este aspecto aún requiere una mayor evidencia científica, ya son varios los trabajos que demuestran una influencia clara del SNC sobre la regulación del ejercicio (39, 40, 56-59). Por lo tanto, aunque esta revisión expresa en muchos casos un punto de vista personal, ésta suposición no debe ser descartada a priori, sino que debe animar a los investigadores ponerla a prueba como hipótesis.
REFERENCIAS BIBLIOGRÁFICAS
1. Breuer HW. Cardiopulmonary exercise tests – proposals for standardization and interpretation. Pneumologie. 2004 Aug;58(8):553-65.
2. Hollmann W. Historical remarks on the development of the aerobic-anaerobic threshold up to 1966. Int J Sports Med. 1985 Jun;6(3):109-16.
3. Taylor HL, Wang Y, Rowell L, Blomqvist G. The Standardization and Interpretation of Submaximal and Maximal Tests of Working Capacity. Pediatrics. 1963 Oct;32:SUPPL 703-22.
4. Calderón FJ, Benito PJ. Ergoespirometría: paradigma del análisis de la respuesta integrada al ejercicio dinámico. Selección. 2001;10(1):21-36.
5. Calderon FJ, Benito PJ, García A. Aplicación práctica de las pruebas de esfuerzo. Selección. 2002;11(4):202-9.
6. Hollmann W. 42 years ago–development of the concepts of ventilatory and lactate threshold. Sports Med. 2001;31(5):315-20.
7. Wasserman K. The anaerobic threshold: definition, physiological significance and identification. Adv Cardiol. 1986;35:1-23.
8. Wasserman K, Beaver WL, Whipp BJ. Gas exchange theory and the lactic acidosis (anaerobic) threshold. Circulation. 1990 Jan;81(1 Suppl):II14-30.
9. Svedahl K, MacIntosh BR. Anaerobic threshold: the concept and methods of measurement. Can J Appl Physiol. 2003 Apr;28(2):299-323.
10. Brooks GA. Anaerobic threshold: review of the concept and directions for future research. Med Sci Sports Exerc. 1985 Feb;17(1):22-34.
11. Wasserman K. Anaerobiosis, lactate, and gas exchange during exercise: the issues. Fed Proc. 1986 Dec;45(13):2904-9.
12. Wasserman K, Whipp BJ, Koyl SN, Beaver WL. Anaerobic threshold and respiratory gas exchange during exercise. J Appl Physiol. 1973 Aug;35(2):236-43.
13. Gaskill SE, Ruby BC, Walker AJ, Sanchez OA, Serfass RC, Leon AS. Validity and reliability of combining three methods to determine ventilatory threshold. Med Sci Sports Exerc. 2001 Nov;33(11):1841-8.
14. Billat VL, Sirvent P, Py G, Koralsztein JP, Mercier J. The concept of maximal lactate steady state: a bridge between biochemistry, physiology and sport science. Sports Med. 2003;33(6):407-26.
15. Iwanaga K, Sakurai M, Minami T, Kato Y, Sairyo K, Kikuchi Y. Is the intracellular pH threshold an anaerobic threshold from the view point of intracellular events?: a brief review. Appl Human Sci. 1996 Mar;15(2):59-65.
16. Loat CE, Rhodes EC. Relationship between the lactate and ventilatory thresholds during prolonged exercise. Sports Med. 1993 Feb;15(2):104-15.
17. Moritani T, Nagata A, Muro M. Electromyographic manifestations of muscular fatigue. Med Sci Sports Exerc. 1982;14(3):198-202.
18. Helal JN, Guezennec CY, Goubel F. The aerobic-anaerobic transition: re-examination of the threshold concept including an electromyographic approach. Eur J Appl Physiol Occup Physiol. 1987;56(6):643-9.
19. Hug F, Laplaud D, Lucia A, Grelot L. A comparison of visual and mathematical detection of the electromyographic threshold during incremental pedaling exercise: a pilot study. J Strength Cond Res. 2006 Aug;20(3):704-8.
20. Jurimae J, von Duvillard SP, Maestu J, Cicchella A, Purge P, Ruosi S, et al. Aerobic-anaerobic transition intensity measured via EMG signals in athletes with different physical activity patterns. Eur J Appl Physiol. 2007 Oct;101(3):341-6.
21. Mateika JH, Duffin J. The ventilation, lactate and electromyographic thresholds during incremental exercise tests in normoxia, hypoxia and hyperoxia. Eur J Appl Physiol Occup Physiol. 1994;69(2):110-8.
22. Matsumoto T, Ito K, Moritani T. The relationship between anaerobic threshold and electromyographic fatigue threshold in college women. Eur J Appl Physiol Occup Physiol. 1991;63(1):1-5.
23. Siciliano G, Pasquali L, Manca ML, Del Corona A, Tessa A, Patrono C, et al. The relationship between anaerobic lactate threshold and plasma catecholamines during incremental exercise in hereditary spastic paraplegia. Funct Neurol. 2003 Apr-Jun;18(2):83-7.
24. Tanabe K, Osada N, Noda K, Yamamoto M, Omiya K, Itoh H, et al. Changes in hemodynamics and catecholamines during single-level exercise at the anaerobic threshold and 120% of the anaerobic threshold in normal subjects. J Cardiol. 1994 Jan-Feb;24(1):61-9.
25. Urhausen A, Weiler B, Coen B, Kindermann W. Plasma catecholamines during endurance exercise of different intensities as related to the individual anaerobic threshold. Eur J Appl Physiol Occup Physiol. 1994;69(1):16-20.
26. Calvo F, Chicharro JL, Bandres F, Lucia A, Perez M, Alvarez J, et al. Anaerobic threshold determination with analysis of salivary amylase. Can J Appl Physiol. 1997 Dec;22(6):553-61.
27. Chicharro JL, Legido JC, Alvarez J, Serratosa L, Bandres F, Gamella C. Saliva electrolytes as a useful tool for anaerobic threshold determination. Eur J Appl Physiol Occup Physiol. 1994;68(3):214-8.
28. Chicharro JL, Lucia A, Perez M, Vaquero AF, Urena R. Saliva composition and exercise. Sports Med. 1998 Jul;26(1):17-27.
29. Segura R, Javierre C, Ventura JL, Lizarraga MA, Campos B, Garrido E. A new approach to the assessment of anaerobic metabolism: measurement of lactate in saliva. Br J Sports Med. 1996 Dec;30(4):305-9.
30. Salminen S, Kontinen A. Effect of exercise on Na and K concentrations in human saliva and serum. J Appl Physiol. 1963 Jul;18:812-4.
31. Conconi F, Ferrari M, Ziglio PG, Droghetti P, Codeca L. Determination of the anaerobic threshold by a noninvasive field in man. Boll Soc Ital Biol Sper. 1980 Dec 15;56(23):2504-10.
32. Conconi F, Grazzi G, Casoni I, Guglielmini C, Borsetto C, Ballarin E, et al. The Conconi test: methodology after 12 years of application. Int J Sports Med. 1996 Oct;17(7):509-19.
33. Debray P, Dey SK. A comparison of the point of deflection from linearity of heart rate and the ventilatory threshold in the determination of the anaerobic threshold in Indian boys. J Physiol Anthropol. 2007 Jan;26(1):31-7.
34. Glass C, Knowlton RG, Sanjabi PB, Sullivan JJ. The effect of exercise induced glycogen depletion on the lactate, ventilatory and electromyographic thresholds. J Sports Med Phys Fitness. 1997 Mar;37(1):32-40.
35. Grazzi G, Casoni I, Mazzoni G, Manfredini F, Uliari S, Conconi F. On the methodology of the Conconi test. Int J Sports Med. 2005 Jun;26(5):397-8; author reply 9.
36. Grazzi G, Casoni I, Mazzoni G, Uliari S, Conconi F. Protocol for the Conconi test and determination of the heart rate deflection point. Physiol Res. 2005;54(4):473-5.
37. Grazzi G, Mazzoni G, Casoni I, Uliari S, Collini G, Heide L, et al. Identification of a Vo2 deflection point coinciding with the heart rate deflection point and ventilatory threshold in cycling. J Strength Cond Res. 2008 Jul;22(4):1116-23.
38. Amann M, Dempsey JA. Locomotor muscle fatigue modifies central motor drive in healthy humans and imposes a limitation to exercise performance. The Journal of physiology. 2008 Jan 1;586(1):161-73.
39. Amann M, Eldridge MW, Lovering AT, Stickland MK, Pegelow DF, Dempsey JA. Arterial oxygenation influences central motor output and exercise performance via effects on peripheral locomotor muscle fatigue in humans. The Journal of physiology. 2006 Sep 15;575(Pt 3):937-52.
40. Amann M, Proctor LT, Sebranek JJ, Eldridge MW, Pegelow DF, Dempsey JA. Somatosensory feedback from the limbs exerts inhibitory influences on central neural drive during whole body endurance exercise J Appl Physiol. 2008;doi:10.1152/japplphysiol.90456.2008.
41. Noakes TD. Physiological models to understand exercise fatigue and the adaptations that predict or enhance athletic performance. Scandinavian journal of medicine & science in sports. 2000 Jun;10(3):123-45.
42. Noakes TD, St Clair Gibson A, Lambert EV. From catastrophe to complexity: a novel model of integrative central neural regulation of effort and fatigue during exercise in humans. British journal of sports medicine. 2004 Aug;38(4):511-4.
43. Noakes TD, St Clair Gibson A, Lambert EV. From catastrophe to complexity: a novel model of integrative central neural regulation of effort and fatigue during exercise in humans: summary and conclusions. British journal of sports medicine. 2005 Feb;39(2):120-4.
44. St Clair Gibson A, Noakes TD. Evidence for complex system integration and dynamic neural regulation of skeletal muscle recruitment during exercise in humans. British journal of sports medicine. 2004 Dec;38(6):797-806.
45. Ulmer HV. Concept of an extracellular regulation of muscular metabolic rate during heavy exercise in humans by psychophysiological feedback. Experientia. 1996 May 15;52(5):416-20.
46. Benito PJ, Peinado AB, Diaz V, Lorenzo I, Calderon FJ. Evolución de los parámetros ergoespirométricos con el entrenamiento en deportistas. Archivos de Medicina del Deporte. 2008;XXIV(6):464-75.
47. Berg K. Endurance training and performance in runners: research limitations and unanswered questions. Sports Med. 2003;33(1):59-73.
48. Zapico AG, Calderon FJ, Benito PJ, Gonzalez CB, Parisi A, Pigozzi F, et al. Evolution of physiological and haematological parameters with training load in elite male road cyclists: a longitudinal study. J Sports Med Phys Fitness. 2007 Jun;47(2):191-6.
49. Zapico AG. Evolución comparada de los parámetros fisiológicos en triatletas y ciclistas de élite, a lo largo de una temporada. Madrid: Universidad Politécnica de Madrid; 2004.
50. Koutedakis Y. Seasonal variation in fitness parameters in competitive athletes. Sports Med. 1995 Jun;19(6):373-92.
51. Pardo FJ. Evolución de los parámetros fisiológicos en ciclistas profesionales a lo largo de una temporada. Madrid: Universidad Politécnica de Madrid; 2001.
52. Svedenhag J, Sjodin B. Physiological characteristics of elite male runners in and off-season. Can J Appl Sport Sci. 1985 Sep;10(3):127-33.
53. Tanaka K, Matsuura Y, Matsuzaka A, Hirakoba K, Kumagai S, Sun SO, et al. A longitudinal assessment of anaerobic threshold and distance-running performance. Med Sci Sports Exerc. 1984 Jun;16(3):278-82.
54. White JA, Quinn G, Al-Dawalibi M, Mulhall J. Seasonal changes in cyclists’ performance. Part I. The British Olympic road race squad. Br J Sports Med. 1982 Mar;16(1):4-12.
55. Yuan Y, Chan KM. A longitudinal study on the ammonia threshold in junior cyclists. Br J Sports Med. 2004 Apr;38(2):115-9.
56. Green AL, Paterson DJ. Identification of neurocircuitry controlling cardiovascular function in humans using functional neurosurgery: implications for exercise control. Exp Physiol. 2008 Sep;93(9):1022-8.
57. Hampson DB, St Clair Gibson A, Lambert MI, Noakes TD. The influence of sensory cues on the perception of exertion during exercise and central regulation of exercise performance. Sports Med. 2001;31(13):935-52.
58. Rauch HG, St Clair Gibson A, Lambert EV, Noakes TD. A signalling role for muscle glycogen in the regulation of pace during prolonged exercise. Br J Sports Med. 2005 Jan;39(1):34-8.
59. Weltan SM, Bosch AN, Dennis SC, Noakes TD. Influence of muscle glycogen content on metabolic regulation. Am J Physiol. 1998 Jan;274(1 Pt 1):E72-82.